
生物学的には, 細胞骨格の微小管内で「レーザーのような」長距離コヒーレント量子現象が発生する可能性がある。この論文では, 私たちが「超放射」および「自己誘導透明性」と呼ぶ現象が生物学的媒体で発生する理論的予測を提示する。微小管の中空コア内に閉じ込められた水分子の電気双極子場と保持されている量子化された電磁放射との相互作用を考慮し, 微小管が非線形コヒーレント光学デバイスの役割を果たすと理論化されている。超放射は, 熱相互作用の特性時間よりもはるかに短い特性時間を持つ特定の量子力学的秩序化現象である。したがって, 微小管内の光信号伝達(および計算)は, 熱雑音と損失の両方から解放される。微小管およびその他の細胞骨格構造のネットワークにおける超放射光計算は, 生体分子認識の基礎と意識の基質を提供する可能性がある。
Keywords: Quantum theory; Quantum coherence; Photon coherence; Neural holography, Microtubules; Spontaneous symmetry breaking; Water molecules; Photon signaling network; Consciousness
脳活動のいくつかの特徴は, 量子物理学との関連を促してきました。1つの特徴は, 意識の物理的相関, つまり多くの実体の多くの状態から構成されるパターンが脳全体に分散していることです。この空間の拡張にもかかわらず, 意識には古典的な手段では説明が難しい「統一性」があります(Marshall, 1989)。非局所的な量子コヒーレンスは, 神経生理学用語で「結合問題」として知られる行き詰まりを打破するために必要な統一性を提供する可能性があります(Crick およびKoch, 1990, Singer, 1993)。
Penrose(1989)が主張したもう1つの特徴は, 多くの脳機能が非アルゴリズム的かつ非計算的であるため, シリアルおよび/またはパラレルの神経処理では説明できないことです。量子コヒーレンス(ペンローズ(1994)によると, 特に微小管における)はこれらの機能を説明できます。意識と量子理論の他のつながりには, 確率的シナプス機能(エクルズ, 1986年, ベックとエクルズ, 1992年), ホログラフィー(プリブラム, 1966年, 1971年, 1991年, ガボール, 1968年), および不確定性原理の一般的な適用(ボーム, 1951年, ボーア, 1958年)などがあります。
1960年代半ば, 光学ホログラフィーの出現は, 脳, 記憶, および知覚の関係を理解する上で新たな出発点となりました。記憶と知覚の側面が比較的広範囲の脳損傷に抵抗することは, 記憶の保存と知覚処理が分散手順であることを示していました。ガボールのホログラフィック方程式のエンジニアリングインスタンスが利用可能になる前は, そのような分散手順がどのようなものになるかを想像することは困難でした。
その後数十年にわたって, コンピュータープログラムはホログラフィーに触発された, ホログラフィック光学手順の本質を構成する並列分散プロセスの連想特性の一部を反映した手法が開発されました。これらの「コネクショニスト」, 「ニューラル ネットワーク」手順により, 多くの知覚および記憶プロセスを in vitro で刺激し, シミュレーションの範囲と限界を調査することが可能になりました。
一方, 修正されていないホログラフィックメタファーは, その拡散関数が無制限(無限)であるため, 脳の処理には不適切なモデルであることが認識されました。神経生理学的実験のデータから, ホログラフィーによってモデル化された皮質機能は, ガウスエンベロープによって制限された正弦波によってより適切に表現されることが示されました。これは, 電話メッセージを大西洋ケーブル経由で送信できる最大効率を測定するために Gabor (1946) によって提唱された数学的定式です。Gabor (1948, 1968) は, ミクロ物理学の単位を説明するためにハイゼンベルクが使用したのと同じ方程式を使用し, そのため, 通信の単位を「情報量子」と呼びました。
したがって, 脳科学が直面する大きな関心事は, 脳による量子情報の処理につながる可能性のあるメカニズムを調査することです。
特定の場の量子論的アプローチは, RicciardiとUmezawa(1967)によって考案されました。彼らは, 脳細胞の構造的構成要素を参照せずに, 量子物理学と記憶の関係を定式化しました。彼らは, 各ニューロンが量子力学的自由度を持つ空間的に分散したシステムを指定し, その物理的特性は場の量子論によって理解できることを強調しました。場の量子論は, 素粒子物理学と凝縮物質物理学の基本プロセスを記述できる量子物理学のフレームワークです(Umezawa, 1993)。RicciardiとUmezawa(1967) は, 記憶は各脳細胞内の空間的に分散した量子力学的自由度の完全な補完の元の動的対称性特性に違反する量子真空状態であると示唆しました。
脳科学における量子論の導入は, 量子宇宙論という新しい分野の最近の出現に似ているかもしれない。宇宙論の基本方程式を「宇宙の波動関数」に作用する量子論的演算子表現として書き直すと, 宇宙は虚数時間で誕生するという解釈が可能になる。ホーキングの提案によれば, 量子トンネル効果によって宇宙は実時間へと進化する。同様に, リッチャルディと梅沢 (1967) が提唱したモデルは, 動的対称性特性に反する真空状態の量子論的概念に依存している。量子論が古典的環境のコンテキスト外で解釈不可能である限り, 量子論と宇宙論の融合は難しいが, 脳科学と量子概念の融合は現在受け入れられている解釈と整合している。外部刺激の記憶は, 真空状態 (つまり, 最低エネルギー状態または基底状態) の順序として刻み込まれる。この秩序だった真空スレートがもはや元の動的対称性特性を示さない場合, 南部-ゴールドストーン定理は, システムの量子力学に参加するゼロエネルギー要件の長距離相関波の生成を示唆します(Ricciardi and Umezawa, 1967)。
このような長距離相関波は, ボーズ量子(「ゴールドストーン粒子」または「質量のある粒子」)として見ることができます。RicciardiとUmezawaは, 脳細胞に量子力学的自由度の全範囲を備えた空間的に分散したシステムの存在を示唆した最初の人物であり, また, 生物学的システムを調査するためのツールとして量子秩序と自発的な対称性の破れを導入しました。少し後に, Fröhlich(1968)は, より明確な物質論的定式化で, 一般的な生物学的細胞に対して同様のアイデアを提案しました。細胞膜のすぐ下の薄い層に焦点を当てて, Fröhlichは, この2次元領域に熱を放出することなくエネルギーを蓄えることができると理論化しました。コヒーレントな双極子伝播波, 細胞膜に隣接した双極子振動活性を持つ生物分子は, 全体的にコヒーレントな動力学モードを発現する可能性があるため, 薄い層は生物学的超伝導媒体, または熱環境から効果的に隔離された生物学的プラズマとして見ることができます。フレーリッヒ波は, フレーリッヒ周波数と呼ばれる1011~1012 s-1の周波数領域で出現すると予測されています。「フレーリッヒ周波数」×プランク定数に等しい大きさで生物細胞に供給されるエネルギーは, 完全に熱化されず, 高度に秩序化された方法で保存される可能性があると結論付けることができます。
生物系におけるフレーリッヒ励起の実験的証拠には, タンパク質におけるGHz範囲のフォノンの観察(Genbergら, 1991年), 鋭い共鳴非熱的!マイクロ波照射による生体細胞への影響(Grundler and Keilmann, 1983), ラット脳におけるGHz誘導性微小管ピノサイトーシスの活性化(Neubauer et al, 1990), および共鳴ラマン法によるフレーリッヒ周波数エネルギーの検出(Genzel et al, 1976: Webb et al., 1977; Webb and Stoneham, 1977: Webb, 1981)。
フレーリッヒ波は, 細胞膜に隣接する薄い層に限定される必要はありません。タンパク質分子の疎水性領域に閉じ込められた水素結合と非局在電子によって維持される双極子振動は, 1 次元媒体を伝播するフロリッヒ波と見なされる集団モードを発現する場合があります。Davydov は, タンパク質内のアルファヘリックスに沿って伝播する双極子孤立波について説明しました。この孤立波伝播モードは、場の量子論では熱化による損失なしにエネルギーと情報を運ぶことが知られており、ダビドフソリトンまたは双極子ソリトンと呼ばれます(Davydov、1979)。
1967年の最初のアイデアから約10年後, 梅沢は2人の同僚である高橋とスチュアートとともに, 各脳細胞内だけでなく脳細胞間でも完全に量子力学的自由度を持つ空間分散システムの洗練された物理モデルを提案しました(Stuartら, 1978, 1979)。脳細胞間に拡張されたシステムは, 量子力学に従う2種類の空間分散自由度で構成されていると考えられています。自発的な対称性の破れの結果として, これら2つのモードは「コルチコン」フィールドと交換ボソンフィールドとして区別され, 量子力学システムとして相互作用します。これは, 非局所的な記憶の保存と想起のプロセスの理論モデルです。最近では, 治部と保江は, コルチコンと交換ボソン場のシステムの物理的に現実的な図を提示し, 新しい場の量子論的枠組みを「量子脳ダイナミクス」(QBD)と呼ぶことを提案しました(Jibu and Yasue, 1992a, b, 19934, b)。彼らは, コルチコンと交換ボソン場を, 細胞骨格と細胞外マトリックスのタンパク質フィラメントと関連する水場に沿って分布する双極振動場として説明しています。マーシャル(1989)は, フレーリッヒの「ポンピングフォノン」が意識の基質である脳内のボーズアインシュタイン凝縮をもたらすと提案しました。
1960年代にリッチャルディと梅沢, フレーリッヒによって始められた脳やその他の生物細胞への量子理論的手法の適用に触発され, 1980年代には他の科学者が量子物理学の微視的観点から脳の機能を捉え始めました。いくつかのグループは, 特に微小管は, 生きた細胞の内部を動的に組織化する糸状タンパク質, すなわち細胞骨格です(HameroffおよびWatt, 1982年: Del Giudice ら, 1983年)。細胞骨格の糸状ポリマー(アクチン, 中間径フィラメントなど)のうち, 微小管は細胞の組織化と情報処理において最も中心的な役割を果たします。微小管は, チューブリンと呼ばれるサブユニットタンパク質二量体の歪んだ六角形の格子で構成され, 直径25nmの中空のシリンダーに自己組織化し, 細胞の活動に適応, 輸送, 制御します。その中空の形状と周期的な格子構造は, いくつかの量子場の側面を示唆しています。Hameroff (1987年) は, 計算セルオートマトンの基礎として, Fröhlichのコヒーレント励起を微小管のチューブリンサブユニットに適用しました。Rasmussenら(1990年)は, このような微小管オートマトンネットワークが学習可能であることを示しました。微小管の外側と内側にある整列した水の層を考慮して, Del Giudiceら(1986)は, 「チューブリン」サブユニットから微小管の円筒形構造が形成されることは, 整列した水による電磁エネルギーの自己集束という概念で理解できると提唱しました。超伝導媒体のマイスナー効果のように, 電磁エネルギーはチューブリンサブユニットが集まるフィラメント領域内に閉じ込められます。Del Giudiceら(1986)は, この自己集束により, 半径15 nmのフィラメントビーム(微小管の内径とまったく同じ)が形成されることを示しました(図1-4を参照)。
図1。急速凍結され, 深くエッチングされたニューロンの微小管(MT)と微小管関連タンパク質(MAP)で重合されたニューロン フィラメントの電子顕微鏡写真。矢印はニューロンフィラメントを指し, 直径が大きい構造は微小管です。シールバー, 100 nm。
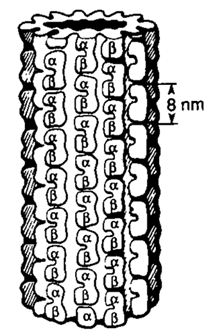
図2。X線結晶構造解析による微小管 (MT) 構造 (Amos および Klug, 1974 年)。チューブリン サブユニットは, α および β モノマーからなる 8 nm の二量体です。
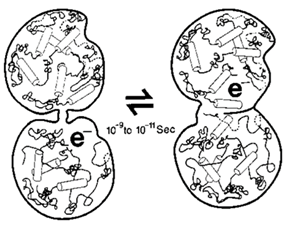
図3。チューブリン二量体タンパク質には, 疎水性領域内の量子イベント(電子移動度, 準粒子フォノン)が 10-9-10-11 秒遷移を伴う構造変化と結合する2つの状態があります。タンパク質内の円筒はαヘリカル領域です
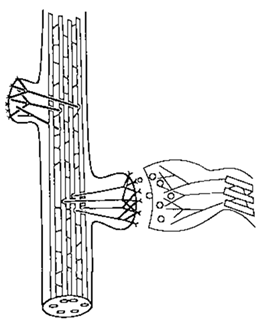
図4。基礎となる細胞骨格構造を示すシナプスの模式図。右: MT-MAPネットワークを持つ軸索末端。シナプス間隙への神経伝達物質小胞(円)の放出に関与するシナプシンフィラメントと接続。左: MT-MAPネットワークを持つ心室樹状突起。細胞骨格構造を持つ2つの樹状突起棘がシナプス膜の受容体と接続。
ハメロフ (1974) は別の観点を提唱し, 微小管が光子, つまり電磁波の量子力学的モードに対する「誘電体導波路」のように機能すると考えられました。実際, 生体組織は非生体物質よりも容易に光を透過します。哺乳類の脳の実験では, 側頭極と海馬で最大の光透過が見られます (Hameroff, 1987)。この証拠から, ハメロフは微小管を光子の導波路と見なしました。さらに, 彼は, 複数の微小管からの, または複数の微小管間のコヒーレント光源の細胞質干渉が, ホログラフィック情報処理メカニズムにつながる可能性があると提唱しました。細胞質ゾルゲル状態に保存されたホログラフィックパターンは, アクチン, アクチン結合タンパク質, 微小柱格子などの細胞骨格のより微細な構成要素のカルシウム誘導アセンブリによって「ハードワイヤード」になる可能性があります。Lechleiter ら(1991) は, 生きた細胞の膜受容体に結合するアセチルコリンによって誘発されるカルシウムのコヒーレントな細胞質螺旋波を画像化した。微小管内の電荷キャリアまたはコンフォメーション波のコヒーレントな発生源は, プリブラムらのホログラフィック脳理論の復活を示唆している (プリブラム, 1966, 1971, 1991, ガボール, 1968, マーゲナウ, 1984, エクルズ, 1986, マーシャル, 1989)。現在の私たちの見解は, コヒーレンスは神経発火レベルのコヒーレンスではなく (またはそれに加えて) 分子, 量子レベルで現れるというものである (シンガー, 1993)。
ハメロフは, 量子物理学の観点から細胞骨格脳活動を理解するための 2 つの重要な概念を非常に一般的な言葉で提案した。それは, 光子の導波路として機能する微小管と, ホログラフィック情報プロセッサとして機能する微小管である。ホログラフィーは, コヒーレントな電磁波または光子の干渉に関連する光学現象です。微小管は光子の導波路とみなすことができ, フロリッヒの理論は微小管におけるコヒーレントな励起を示唆しています。さらに, 微小管の周期的な格子構造は, 光子が通過できる周期的に配列された「スリット」(二量体間の空間)を提供する可能性があります。光学ホログラフィーに必要なコヒーレントな光子の生成または放出の証拠は, 微小管または微視的な生物学的構造では見つかっていません。ただし, 自己収束光子および自己捕捉された非熱化波実体は検出が困難なはずです(セクション 4 を参照)。いずれにせよ, 水レーザー(Del Giudice 他, 1988 年)を除いて, 物理学の観点からこれらの概念の徹底的な理論的調査は行われていません。
本論文では, 超放射および自己誘導透明性と呼ばれる協調的な量子ダイナミクスを生み出す秩序立った体系的な特性を微小管が備えていることを示唆する新しい量子理論的枠組みを提案します。具体的には, 水分子の量子力学システムと中空の微小管コア内に閉じ込められた量子化された電磁場は, 超放射と呼ばれる特定の集合的ダイナミクスを発現することができ, これにより微小管は, 非干渉性, 熱的, 無秩序な分子, 電磁気, または原子エネルギーを微小管内でコヒーレントな光子に変換することができます。さらに, 超放射によって生成されたこのようなコヒーレントな光子は, あたかもその中の光学媒体が伝播する光子自体によって「透明」になったかのように, 微小管の内部中空コアに沿って完全に浸透することも示されます。これは, 自己誘導透明性と呼ばれる量子理論現象です。
細胞骨格微小管の超放射と自己誘導透明性は, 「光学的」神経ホログラフィーにつながる可能性があります。ニューロン(および他の細胞)には, ホログラフィック記憶操作を備えた『2001年宇宙の旅』の架空だが有名なHAL 9000のような, 膨大な容量を持つ微小なコヒーレント光スーパーコンピュータが含まれている可能性があります(Clarke, 1968年)。
この論文は, 物理学者と脳科学者の長距離協力の結果であり, いくつかの副産物を生み出しています。
脳細胞や一般的な生物細胞の細胞骨格構造にある典型的な微小管を見てみましょう。これは直径約 25 nm の中空の円筒で, その壁はタンパク質サブユニット (「チューブリン」) の重合配列です。その長さは数十ナノメートルからマイクロメートルの範囲で, 大型動物の神経軸索ではさらに数メートルになることもあります。物理的な定式化を簡素化するために, 微小管を半径 \(r_{MT}\), 長さ \(l_{MT}\) の中空の円筒と見なします。実際の値は \(r_{MT}≒12, l_{MT}≒10^2-10^3\) (ナノメートル単位)になります。微小管シリンダー内の空間領域を \(V\) で表し, この空間領域 \(V\) で発生する量子力学ダイナミクスに議論を限定します。
原点 \(O\) がエンドキャップの中心と一致するように, 微小管シリンダーの2つの端の1つに平行に \(xy\) 平面が取り付けられた直交座標系 \(O_{xyz}\) を導入します。\(z\) 軸は, 微小管シリンダーの縦方向の中心軸に沿って自然に存在します。次に, 領域 \(V\) 内の任意の位置を, その座標 \(r = (x,y,z)\) を与えることでラベル付けできます。
微小管内の空間領域Vは空ではありませんが, 水分子で満たされている可能性があります。もちろん, 他の分子が存在する可能性もありますが, その数は比較的少ないです。水以外の分子の存在を無視できる理想的なケースを検討します。ただし, 以下では, 麻酔分子などの水分子に不純物がある, より現実的なケースを検討します。おそらく, 微小管シリンダー内に閉じ込められた水の密度はほぼ一定のままです。したがって, 領域 \(V\) 内の水分子の総数を \(N\) と固定できます。
典型的な水分子, たとえば \(j\) 番目の水分子を見てみましょう。ここで, 1 から \(N\) までの \(j\) は, 問題の \(V\) 個の水分子に付けられた架空の番号を表します。その位置は, 座標 \(r_j = (x_j, y_j, z_j)\) で与えられます。物理的な観点から, 水分子は一定の電気双極子を持つため, 電気双極子モーメントを持つ量子力学的こまとして見ることができます。水分子の平均慣性モーメントと電気双極子モーメントは, それぞれ \(l = 2m_pd^2 (d \approx 0.82 Å), μ= 2e_pP (P \approx 0.2 Å)\) と推定されます。ここで, \(m_p\) は陽子の質量, \(e_p\) は陽子の電荷を表します。
電気双極子モーメント \(μ\) により, 水分子は空間領域 \(V\) の量子化電磁場と強く相互作用します。水分子は量子力学のこまとして多くのエネルギー固有状態を持ち, さまざまな値の量子化電磁場間でエネルギーを交換できますが, ここでは, 2 つの主要なエネルギー固有状態のみがエネルギー交換に参加する最も可能性の高いケースに限定して説明します。これは, レーザー理論で原子と量子化電磁場間のエネルギー交換を記述する際の従来の 2 レベル近似と一致します。
すると, \(j\) 番目の水分子の量子ダイナミクスは, スピン変数 \(s^j = 1/2σ\) で適切に記述できることがすぐにわかります。ここで, \(σ = (σ_x, σ_y, σ_z)\) であり, \(σ_x\) は, スピン 1/2 の角運動量の 3 つの成分を表すパウリスピン行列です。水分子の 2 つの主要なエネルギー固有状態間のエネルギー差を \(\epsilon\) とします。その実際の値は \(\epsilon \approx 200 cm^{-1}\) です (Franks, 1972)。次に, \(j\) 番目の水分子の量子力学を支配するハミルトニアンは \(\epsilon s_z^j\) で与えられ, \(NV\) 水分子の全ハミルトニアンは \[ H_{WM}=\epsilon \sum_{j=1}^N s_z^j \tag{1} \] 前者のハミルトニアンの 2 つのエネルギー固有値は, エネルギー差 \(\epsilon\) を持つ 2 つの主要なエネルギー固有状態のみが考慮されているという事実を反映して, \(-1/2\epsilon\) と \(1/2\epsilon\) です。
さて, 空間領域 \(V\) の量子化された電磁場を考えてみましょう。量子化された電磁場は, 電場演算子 \(\mathbf{E }= \mathbf{E}(\mathbf{r}, t)\) で記述すると便利です。簡単にするために, 電場が直線偏光していると仮定して, \(\mathbf{E} = \mathbf{e}E\) を得ます。ここで, \(\mathbf{e}\) は直線偏光の方向を指す単位長さの定数ベクトルです。すると, 問題の量子化された電磁場は, 通常のハミルトニアンによって支配されるスカラー電場 \(E = E(\mathbf{r}, t)\) によってうまく記述されるようになります。 \[ H_{EM}=\frac{1}{2} \int_V E^3d^3r \tag{2} \]
次に, 量子化された電磁場と水分子全体との間の相互作用を考えます。この相互作用によって, 水分子は光子の生成と消滅という形でエネルギーを交換できます。電場演算子を正と負の周波数部分に分けてみましょう \[ E=E^+ + E^- \tag{3} \] すると, 量子化された電磁場と水分子全体の相互作用ハミルトニアンは \[ H_I=-μ\sum_{j=1}^N [E^- (\mathbf{r}^j,t) s_-^j+s_+^j E^+(\mathbf{r}^j,t)] \tag{4} \] ここで \[ s_±^j=s_x^j±is_y^j \tag{5} \] 電磁場の量子力学, 水分子の双極子振動場, およびそれらの相互作用を支配する全ハミルトニアンは \[ H=H_{EM}+H_{WM}+H_I \tag{6} \] 微小管円筒内部の空間領域Vは電磁波の空洞とみなせるため、電界演算子 \(E = E^++E^-\) の通常モード展開を導入すると便利であり、 \[ E^±(\mathbf{r},t)=\sum_k E_k^± (t) e^{±i(\mathbf{k}\cdot \mathbf{r}-ω_k t)} \tag{7} \] ここで, ωk は波動ベクトル \(\mathbf{k}\) で作成された法線の固有角周波数を表します。我々が主に関心があるのは, 水分子間の秩序だった集団的行動とキャビティ領域 \(V\) 内の量子化された電磁場です。そこで, 水分子の集団的動的変数 \(S_k^±(t)\) と \(S\) を \[ S_k^±(t)=\sum_{j=1}^N s_±^j(t) e^{±i(\mathbf{k} \cdot \mathbf{r}^j-ω_k t)} \tag{8} \] および \[ S=\sum_{j=1}^N s_z^j \tag{9} \] で導入します。すると, 全ハミルトニアン(式6) は \[ H=H_{EM}+\epsilon S-μ \sum_{k} (E_k^- S_k^-+S_k^+ E_k^+) \tag{10} \]
ここで注目すべきは, \(N\) 個の水分子と微小管シリンダー内の領域 \(V\) 内の量子化された電磁場のシステムに対するこの全ハミルトニアンは, レーザーシステムに対する Dicke のハミルトニアンだけでなく, 量子脳ダイナミクスに対する Stuart らのハミルトニアン (Dicke, 1954 年, Stuart ら, 1979年) と本質的に同じ形式であるということです。したがって, 脳細胞の細胞骨格構造内の各微小管は, QBD における記憶の書き込みと想起のメカニズムだけでなく, レーザーのようなコヒーレントな光学活性も示すことが期待できます。前者は, Jibu と Yasue (1992a, 1993a, b)が水分子とタンパク質フィラメントの観点から QBD の物理的描像を展開したため, それほど驚くべきことではありません。したがって, 本稿ではこれについては説明しません。一方, 後者は, 従来のものとは大幅に異なる, 脳機能の基本的プロセスのまったく新しい描像を開く可能性があるため, 非常に驚くべきようです。つまり, 神経細胞間の膜貫通イオン拡散の観点から神経インパルスの通常の経路伝導に加えて, 脳システムは, レーザー, 光ファイバー, その他の光学デバイスを使用した光コンピューターと同様に, 微小管内のコヒーレントな光子放出と転送の観点から, 別のはるかに微視的で精巧な基本的物理プロセスを使用している可能性があります。この驚くべき可能性について, 本論文ではずっと議論していきます。
しかし, たとえ微小管が水分子と電磁場に関して, 典型的なレーザーシステムのハミルトニアンによって支配されるのと同じ量子力学的挙動を示すとしても, 脳細胞内でレーザーデバイスを実現する可能性を探すのは困難です。これは, 最初に, 特定の非コヒーレントだが高強度の光によって, 大部分の水分子をより高いエネルギーの固有状態にポンピングする必要があるためです。たとえば, 実際の人工レーザーデバイスでは, 最初のポンピングはキセノンフラッシュランプによるものです。言い換えれば, レーザーシステムは, 何らかのポンピングメカニズムなしではコヒーレント光子を放出できません。脳内にそのようなポンピング光が存在することはほとんど期待できません。したがって, ポンピング光なしでコヒーレント光子放出の異なるメカニズムにつながる合計ハミルトニアン(式10)を持つ別の可能性を探さざるを得ません。
幸いなことに, 全ハミルトニアン(式10)の形式を物理的に検査すると, 基底状態では明らかではない動的対称性特性が表れることが明らかになり, その結果生じる量子ダイナミクスには, 自発的な対称性の破れによる特定の長距離秩序生成現象が含まれることが知られています(Stuartら, 1979)。この長距離秩序の空間次元, つまり, コヒーレンス長 \(l_c\) は, エネルギー差 \(e\) に反比例すると推定され, \(l_c \approx\) 数百マイクロメートルになります。これらの長距離秩序生成現象の中には, 微小管シリンダー \(V\) 内の大多数の水分子の集団ダイナミクスが, ポンピング光なしで光子の協調的な自発放出を引き起こす特定の現象が見つかるかもしれません。微小管シリンダーの壁を形成するタンパク質サブユニット(チューブリンなど)の重合配列のマクロな熱力学による水分子間の非コヒーレントで無秩序なエネルギー分布は, コヒーレントで秩序立ったダイナミクスに集合的に集められ, コヒーレント光子を協調的に放出する準備が整います。ポンピング光なしでコヒーレント光子を放出するこのレーザーのようなプロセスは, Dicke(1954) によって初めて導入され, 超放射と呼ばれました。
微小管シリンダー \(V\) の超放射を全ハミルトニアン(式10)から調べましょう。簡単にするために, 特定の波数ベクトル(たとえば k0)を持つ1つの標準モードのみが, 2つの主要なエネルギー固有状態間のエネルギー差εに共鳴する適切な角周波数ωk0を持つと仮定します。つまり, 次のようになります。 \[ \epsilon = \hbar \omega_{k_0} \tag{11} \] 他のすべての標準モードは無視されます。従来のレーザー理論では, これはシングルモードレーザーとして知られています。以下で分析するものは, したがって, 微小管内の単一モードの超放射として考えることができます。 波数ベクトルk0を持つ1つの通常モードしかないため, 動的変数の波数ベクトル インデックスをすべて省略できます。すると, 合計ハミルトニアン (式 10) は次のようになります。 \[ H=H_{EM}+\epsilon S-μ(E^- S^-+S^+ E^+) \tag{12} \] 水分子の 3 つの集団動的変数 \(S\) と \(S^±\), および量子化された電磁場の 2 つの変数 \(E^±\) に対応するハイゼンベルクの運動方程式は, 次の式で与えられます。 \[ \begin{align} \frac{dS}{dt} &=-i \frac{μ}{\hbar} (E^- S^--S^+ E^+) \tag{13} \\ \\ \frac{dS^±}{dt} &=±i \frac{2μ}{\hbar} SE^∓±i \frac{\epsilon}{\hbar}S^± \tag{14} \end{align} \] および \[ \frac{dE^±}{dx} =±i \frac{2π\epsilon μ}{\hbar V} S^∓ \tag{15} \] 微小管シリンダーの長さが短いため (\(l_{MT} \approx 10^2-10^3 nm\)), \(z\) 軸方向に微小管シリンダーに沿って伝播するパルスモードは, キャビティ領域 \(V\) に, 短い通過時間 \(t_{MT}=l_{MT}/c\) だけ留まります (\(c\) は光速を表します)。このパルスモードの通過時間は, 無秩序な環境による熱相互作用の特性時間よりもはるかに短いため, この単一モード超放射における水分子と量子化電磁場のシステムは熱損失がなく, ハイゼンベルクの運動方程式(式13-15)によってよく説明される閉鎖系と見なすことができます。さらに, 量子化電磁場の動的変数 \(E^±\) の時間微分は, 微小管の縦軸に沿って伝播するパルスモードの場合, \(E^±/l_{MT}\) で近似できます。式15から次の式が得られます \[ E^±=±i \frac{2π\epsilon μl_{MT}}{\hbar V} S^∓ \tag{16} \] これは, 微小管シリンダーキャビティ内の量子化電磁場のパルスモードが, 微小管内の水分子の集団ダイナミクスに従うことを意味します。言い換えると, 自発的な対称性の破れにより水分子のダイナミクスに長距離秩序を持つ集団モードが生成されると, 量子化電磁場のパルスモード, つまり光子のコヒーレント放出が続きます。これが超放射のメカニズムです。
最後の疑問は, このような集団モードが, 非コヒーレントで無秩序な初期条件から始まる水分子のダイナミクスで実現できるかどうかです。水分子のこのような非コヒーレントで無秩序な初期の動的構成は, 水分子とチューブリンの配座ダイナミクスの熱無秩序状態との相互作用によるものであることに注意してください。この集団モードの開始は, 水分子の集団変数に対する3つのハイゼンベルク方程式(式13と14)を式16に置き換えて書き直すことで確認できます。つまり, 次の式が得られます。 \[ \frac{dS^±}{dt} =βSS^±±i\epsilon S^± \tag{17} \] および \[ \frac{dS}{dt}=-βS^+ S^- \tag{18} \] ここで \[ β=\frac{4π\epsilon μ^2 l_{MT}}{\hbar^2 V} \] これらは, 特定の交換関係に従う非交換演算子SとS±の結合非線形微分方程式であり, その解を見つけるのはそれほど簡単ではありません。しかし, これらの方程式を古典的(つまり可換な)動的変数に対する通常の結合非線形微分方程式とみなすと, 非コヒーレントで無秩序な初期条件に関して特別な解を見つけることができます。これらは, 超放射の量子力学系の半古典的近似として知られています(Agarwal, 1971)。簡単な計算により, 超放射による微小管シリンダー内のコヒーレント光子放出の強度は, この近似で次のように表すことができます \[ I=\frac{\hbar^2}{4t_R μ)^2} sech^2 \frac{t-t_0}{2t_R} \tag{19} \] ここで \[ t_R=\frac{c\hbar^2 V}{4πμ^2 \epsilon Nl_{MT}} \] および \(t_0 = t_R\ln2N\) は, それぞれ超放射の寿命と遅延を表します。超放射の強度は \(N^2\) に比例し, 遅延時間は \(N\) に反比例することに注目してください。これらの事実は, \(N\) 水分子を含む長距離秩序生成プロセスの特徴です (図 5 を参照)。
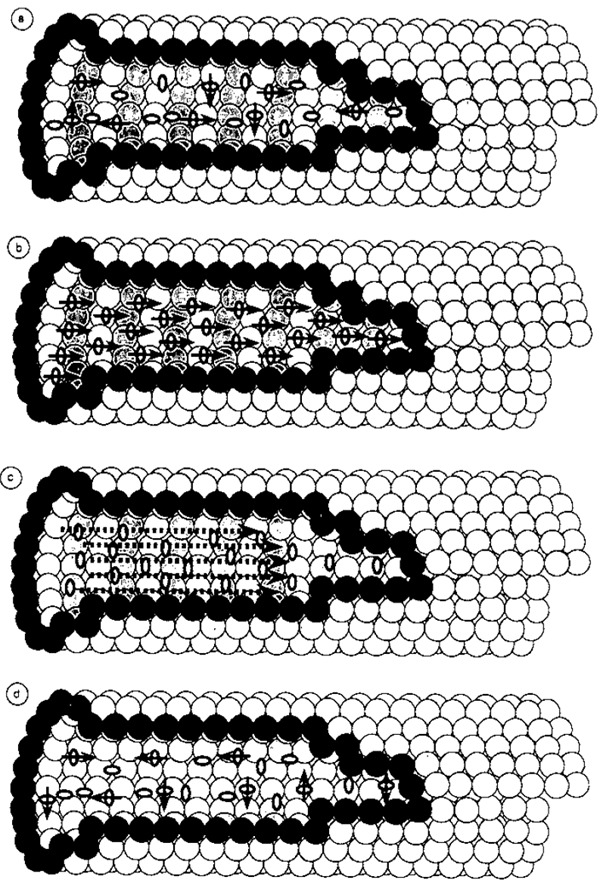
図 5。微小管における超放射プロセスの概略図。矢印のない楕円は, 最低回転エネルギー状態にある水分子を表します。矢印のある楕円は, 最初の励起回転エネルギー状態にある水分子を表します。プロセスは循環的です (a → b → c → d → a → b)。 (a) 微小管内の水分子システムの初期状態。チューブリンの熱変動によるエネルギー増加により, 最初の励起回転エネルギー状態にある水分子の数が増加します。 (b) 回転励起状態にある水分子システムの集団モード。長距離コヒーレンスは, 自発的な対称性の破れによって微小管内で達成されます。(c) 回転励起状態にある水分子のシステムの集団モードは, 集団的にエネルギーを失い, 微小管内の量子化された電磁場にコヒーレント光子を生成します。(d) 超放射によって最初の励起回転エネルギーを失った水分子は, チューブリンの熱変動から再びエネルギーを獲得し始め, 水分子システムは初期状態 (a) を回復します。
私たちは, 水分子の量子集団ダイナミクスと微小管シリンダー内の量子化された電磁場が, 超放射の長距離協力現象を発現することを発見しました。この現象では, 微小管シリンダーの壁を形成するタンパク質分子のマクロな熱ダイナミクスによる非コヒーレントで無秩序な摂動によって, 水分子の集団励起が誘発される可能性があります。この事実は, 脳細胞, すなわちニューロンとアストロサイトの細胞骨格構造における各微小管が, 水分子とタンパク質分子のマクロな無秩序なダイナミクスを, 水分子の長距離秩序のあるダイナミクスと, コヒーレント光子のパルスモード放出を伴う量子化された電磁場に変換する超放射デバイスとして, 脳機能の光情報処理レジメンにおいて重要な役割を果たす可能性があることを保証します。言い換えれば, 各微小管は, 各脳細胞の細胞質内の高密度の微視的光コンピューティングネットワーク内のコヒーレント光エンコーダです(そのようなネットワークが実際の細胞質構造で実現されている場合)。この最後の点はまったく明らかではなく, 次のセクションで議論する価値があります。
微小管の超放射によるコヒーレント光子放出の観点から, 基本的な脳機能のまったく新しいメカニズムの可能性を示しました。レーザーとは異なり, 超放射は, 熱相互作用よりはるかに短い特性時間を持つ特定の量子力学的秩序化プロセスです。したがって, 微小管は, (1) 膜貫通イオン拡散と熱的に摂動された分子振動の観点から見た古典的で無秩序で非コヒーレントな神経ダイナミクスの従来のマクロシステムと, (2) 熱雑音と損失のない秩序立ったコヒーレントな量子ダイナミクスのこれまで知られていないミクロな光コンピューティングネットワークシステムとの間の物理的インターフェイスを提供する理想的な光エンコーダーであると考えられます。Feynman (1985) が提案したように, 光コンピューティングネットワークは, 超伝導コンピュータなどの多くの可能性の中で最も実現可能な量子力学的コンピュータです。ただし, 超放射によって微小管シリンダー内に生成されたパルスモードのコヒーレント光子が, その長距離コヒーレンスを維持したまま安全に伝送できるかどうかは明らかではありません。超放射によって放出されたコヒーレント光子でさえ, ノイズの多い熱環境のために, コヒーレンスと長距離秩序をすぐに失う可能性が高いと思われます。いずれにせよ, 脳細胞内のまだ知られていない微視的光コンピューティング ネットワークで光子のコヒーレント伝送を維持するには, 何らかの量子力学的メカニズムが絶対に必要です。このセクションでは, 脳細胞の細胞骨格構造内の微小管が, このようなメカニズムを提供するものであることを示します。
パルスモードコヒーレント光子が超放射によって微小管円筒の小さなセグメントに生成されたと仮定します。次に, それらの光子は微小管円筒の縦軸, つまり z 軸に沿って伝播します。微小管円筒内の領域Vが真空に維持されている場合, 光子は導波管に沿って移動するのと同じようにその領域を通過します。ただし, その領域は水分子で満たされており, パルスモードコヒーレント光子が吸収やコヒーレンスの損失なしにその領域を安全に透過できるかどうかはまったく明らかではありません。
パルスモードコヒーレント光子が微小管円筒内の領域に沿って透過することを説明するためには, 水分子の長距離秩序ダイナミクスが量子力学によって記述され, 電磁場が古典的に記述される半古典的近似で作業するのが便利と思われます。この近似は, パルスモードのコヒーレント光子の強度が大きい場合に信頼性が高くなり, これは超放射放射の場合に実際に当てはまります (Feynman ら, 1957)。
スカラー電場 \(E= E(z, t)\) の Maxwell 方程式を考えてみましょう。これは, 微小管シリンダー内の水分子の集団的動的変数 \(S^±\) と結合した \(z\) 軸に沿って伝播するパルスモードのコヒーレント光子を表します。 \[ \frac{∂E^±}{∂z}+\frac{∂E^±}{∂t}=∓i \frac{2π\epsilon μ}{\hbar V} S^∓ \tag{20} \] この方程式は, 集団的動的変数が変数 \(S^±\) はゆっくりと変化します。つまり, \[ \frac{∂S^±}{∂t} \ll iωS^± \tag{21} \] 次に, このマクスウェル方程式とハイゼンベルクの運動方程式(式13と14)における量子力学的変数の期待値を取り, 水分子に関する変数の期待値をすべて消去し, スカラー電場の新しい変数を導入して, \[ θ^±(z,t)=\frac{2μ}{\hbar} \int_{-\infty}^t E^± (z,u)du \tag{22} \] によって導入すると, 微小管シリンダー内の領域 \(V\) の電磁場の動的方程式が得られます \[ \frac{∂^2 θ^±}{∂τ∂ζ}=-\sin θ^± \tag{23} \] ここで \[ τ=\sqrt{\frac{2π\epsilon μ^2 N}{\hbar^2 V}}\left(t-\frac{z}{c}\right) \] および \[ ζ=\sqrt{\frac{2π\epsilon μ^2 N}{\hbar^2 V}}\frac{z}{c} \] これは, サインゴードン方程式と呼ばれる典型的な非線形偏微分方程式であり, ソリトン方程式の逆散乱法によっていくつかの正確な解が得られます(Ablowitzら, 1974)。
サインゴードン方程式 (式 23) の最も興味深い解は, 領域 \(V\) のスカラー電場Eの時間発展の明示的な形式を生み出します微小管シリンダー内部 \[ E=\sqrt{\frac{2π\epsilon Nv_0}{V(c-v_0)}} sech\sqrt{\frac{2π\epsilon μ^2 Nv_0}{\hbar^2 V(c-v_0)}}\left(t-\frac{z}{v_0}\right) \tag{24} \] これはソリトン解に他ならず, パルスモード光子が, 真空中の光速 \(c\) より遅い一定の速度 \(v_0\) で水分子で満たされた微小管シリンダーの誘電体導波路に沿って伝播することを示しています。ソリトン解のパルス形状は, サインゴードン方程式の非線形性により変化しないことを確認することが重要です。
微小管は誘電体導波路の役割を果たしており, パルスモードのコヒーレント光子は, 完全に透明であるかのように微小管を伝播することがわかっています。この現象は自己誘導透明性と呼ばれ, 量子光学における典型的な非線形効果として知られています(McCall and Hahn, 1967)。微小管は, 熱雑音や損失のない, パルスモード光子の完全に透明な経路として使用するのに最適な微視的光学デバイスである可能性があります。超放射と組み合わせると, この微小管の自己誘導透明性から, 脳は本質的に, 脳細胞の細胞骨格構造にある微小管の微細で精巧な光コンピューティングネットワークの密集した集合体であると結論付けることができます。各微小管におけるコヒーレントな光子の放出と転送は, 量子力学における長距離秩序化現象に特徴的な超放射と自己誘導透明性によって保証されます。
超放射と自己誘導透明性はどちらも, 単一細胞内および多数の細胞間の細胞骨格構造における複数の微小管の協調的な物理的活動に不可欠である可能性があります。したがって, 一般的な生物システム, 特に脳は, 量子レベルのホログラフィック情報システムを利用している可能性があります(Hameroff, 1987年, Pribram, 1991年)。
私たちは, 微小管とそれに関連する水における光子の量子コヒーレンスの発生に関する理論的枠組みを開発しました。微小管と細胞骨格が「リアルタイム」の細胞機能の編成と制御に根本的に関与していることは, そのような量子コヒーレンスが存在する場合, 細胞通信, シグナル伝達, 情報処理において重要な役割を果たす可能性があることを示唆しています (Conrad, 1988)。
微小管における量子コヒーレンスには, いくつかの側面が考えられます。
導波管として働く円筒形微小管の中空コア内の整列した水で発生する超放射と自己誘導透過性は, コヒーレント光子をもたらす。このコヒーレンスは, 数百マイクロメートルにわたって空間的に分布する微小管間の状態の重ね合わせが可能と推定され, さらに, 他の方向に数百マイクロメートル離れた他の微小管と重ね合わせられるなど, 広範囲にわたる微小管ダイナミクスの結合を説明できる可能性がある。これは, 思考と意識の統一を説明できる可能性がある。たとえば, Insinna (1992) は, ユングのシンクロニシティ現象を微小管の量子コヒーレンスに起因するものとしている。
ホログラフィック情報表現につながるコヒーレント干渉は, 量子光学効果から生じる。細胞骨格レベルでは, カルシウム誘導ゾルゲル変化によってアクチンやその他の細胞骨格フィラメントの重合が起こり, 短期記憶のホログラフィック表現が生じる可能性がある(Hameroff, 1987)。ホログラフィックパラダイムは, 複数のスケールにわたるコヒーレンスとホログラフィック干渉を示唆している(Pribram, 1991)。より「マクロ」レベルでは, 40 Hzの範囲で広く分散したニューロン間のニューロン発火のコヒーレンスは, コヒーレントに発火するニューロン内の微小管の量子コヒーレンスに関連している可能性がある(Singer, 1993)。
個々の微小管サブユニットにおける量子イベントの結果として, 音響コンフォメーション振動が細胞骨格ネットワーク, 膜, 細胞外マトリックスを介して信号として伝播する可能性がある。このような提案された信号(セルオートマトン理論の文脈で)は, 理論的には学習, 論理, 適応が可能であることが示されています(Rasmussen et al., 1990; Lahoz-Beltra et al., 1993)。
私たちは, 細胞骨格とそれに関連する水に由来し, 他の生体構造に協力的に浸透する複雑なエネルギー/情報フィールドについて説明しました。「統一場理論」は生物学と生物物理学において長く失敗した歴史があり, 「生気論」という軽蔑的な用語を暗示しています。私たちは, 生物学的情報, コミュニケーション, および外部世界の表現の媒体として, 微小管由来のゴールドストーン粒子に基づく新生気論的量子コヒーレントホログラフィックフィールドを導入します。最大の「脳全体」スケールでは, これが意識を構成するはずです。
ATPとGTPの加水分解とタンパク質のリン酸化による生化学的エネルギーによって駆動される微小管の量子コヒーレンスと自己フォーカスは, 全身麻酔に敏感であるはずです。麻酔ガス分子は, タンパク質の疎水性領域での弱いファンデルワールス結合によって意識を可逆的に抑制します(FranksとLieb, 1982)。微小管の量子光学コヒーレンスが意識に不可欠である場合, 麻酔薬は何らかの方法でそれを阻害する必要があります。考えられる方法の 1 つは, 細胞骨格のダイナミクスを外部環境から切り離して隔離できる膜阻害です。あるいは, 麻酔薬が細胞骨格の量子コヒーレンスを直接変更する可能性があります。全身麻酔薬は微小管に結合し, 十分な濃度になると脱重合を引き起こします(Allison と Nunn, 1968)。意識を消失させるのに十分な麻酔濃度では, チューブリンのコンフォメーション ダイナミクスの阻害が大きな影響を及ぼす可能性があります。Wulfと Featherstone(1957)は, タンパク質の疎水性領域内での麻酔薬の結合が, タンパク質表面でのタンパク質と水の結合を変化させることを示した。したがって, チューブリンの立体配座ダイナミクスと協同的な水結合に対するわずかな麻酔効果は, 量子コヒーレンスに重大な影響を及ぼす。麻酔は圧力によって解除されることも知られている(Halsey, 1976)。非線形光学媒体を凝縮することにより, 圧力は量子協同コヒーレンス現象を増加させる。したがって, 私たちのモデルは全身麻酔のメカニズムと一致している。
この論文では, 細胞骨格微小管における量子光コヒーレンスの理論的枠組みを提示する。具体的には, 水分子の量子力学システムと中空微小管コア内に閉じ込められた量子化された電磁場は, 超放射と呼ばれる特定の集団ダイナミクスを示し, それによって微小管内でコヒーレント光子が生成される。さらに, 超放射によって生成されたこのようなコヒーレント光子は, あたかもその内部の光学媒体が伝播する光子自体によって透明になったかのように, 微小管の内部中空コアに沿って完全に浸透します。この量子効果は, 自己誘導透明性と呼ばれます。このような効果は, 細胞シグナル伝達, ホログラフィック情報処理, 意識の統一を説明することができます。
著者一同、H. 梅沢教授、Y. 高橋教授、L. M. Ricciardi 教授、C.I.J.M.Stuart教授に、生体物質の量子論的枠組みを提供してくださったことに感謝申し上げます。C.I.J.M.Stuart 教授のご逝去を知り、大変悲しく思っております。私たちは彼に多大な恩義を感じており、深く哀悼の意を表します。Stuart R・Hameroff 研究は、NSF助成金番号DMS-9114503の助成を受けています。